R.F. Miller and J.A. Rose
Originally published in:
Great Basin Naturalist
1995, Volume 55, No. 1, pages 37-45
- Abstract
- Introduction
- Study Area
- Plot Layout
- Measurements
- Statistical Analysis
- Results
- Discussion
- Conclusion
- Literature Cited
Abstract
The chronology of Juniperus occidentalis (western juniper) expansion in eastern Oregon, the
effect of plant canopy and interspace on J. occidentalis seedling establishment and growth rates,
and the age of J. occidentalis maximum reproductive potential were determined. Measurements
were recorded in twenty-two 0.4-ha plots established in sagebrush-grassland communities and six
0.1-ha plots in Populus tremulodes (quaking aspen) communities. J. occidentalis began
increasing during the 1880s in stands containing trees > 130 yr old. Relatively steady
establishment ensued into the 1950s and then began to progress at a geometric rate in the 1960s.
J. occidentalis encroachment into aspen stands began between 1910 and 1920. The largest
proportion of juvenile trees established beneath Artemisia species in sagebrush- grassland
communities. J. occidentalis trees appeared to reach full reproductive potential at >50 yr of age.
The ratio of male:female trees increased from 1.7 in scattered J. occidentalis stands to 3.8 in
closed stands. The initiation of J. occidentalis encroachment during the late 1800s coincides with
optimal climatic conditions for Juniperus berry production and establishment, reduced fire-return
intervals, and heavy livestock grazing. The accelerated increase in J. occidentalis expansion since
1960 may be due to the continued absence of fire, abundant woody plant cover, and the large
increase in J. occidentalis seed production.
Return to Contents
Introduction
One of the most pronounced plant community changes in the 20th century has occurred in the
juniper and pinyon-juniper woodlands, a major vegetation type characterizing the Intermountain
Region. These woodlands, sometimes described as pygmy forests, currently occupy 17 million ha
throughout this region (West 1988). Juniperus occidentalis ssp. occidentalis Hook. (western
juniper) is considered the Northwest representative of the pinyon-juniper zone in the
Intermountain Region (Franklin and Dyrness 1973) and occupies over 1 million ha (Dealy et al.
1978) in eastern Oregon, southwestern Idaho, and northeastern California (Cronquist et al. 1972).
This subspecies of J. occidentalis is found primarily north of the polar front gradient (Neilson
1987; parallel to the Oregon and Nevada border, latitude 42°) where temperatures are cooler,
summer precipitation decreases, and winter precipitation increases (Mitchell 1976).
Relict juniper woodlands, tree-age class distribution, fire scars, and historical documents indicate
presettlement pinyon-juniper and juniper woodlands were usually open, savannah-like (Nichol
1937, West 1988), or confined to rocky surfaces or ridges (Cottam and Stewart 1940, Barney and
Frishknecht 1974, Hopkins 1979, Johnson and Simon 1987). J. occidentalis began increasing in
both density and distribution in the late 1800s (Burkhardt and Tisdale 1976, Young and Evans
1981, Eddleman 1987), invading Artemisia tridentata subsp. vaseyana (mountain big sagebrush),
Artemisia arbuscula (low sagebrush), Populus tremuloides (quaking aspen), and riparian
communities. Although J. occidentalis is long lived (Vasek 1966, Lanner 1984), less than 3% of
the woodlands in Oregon are characterized by trees > 100 years old (USDI-BLM 1990). In 1825,
Ogden observed only occasional J. occidentalis (reported as cedars) growing on hillsides while
traveling through the Crooked River drainage in central Oregon (Rich et al. 1950). Today, these
hillsides are covered by dense J. occidentalis woodlands. In a nearby area J.W. Meldrum's 1870
survey notes describe a gently rolling landscape covered with an abundance of perennial
bunchgrasses and a wide scattering of J. occidentalis trees (Caraher 1977). Today, J. occidentalis
densities on this site range between 125 and 250 ha-1. In Silver Lake, Oregon, J. occidentalis
density increased from 62 ha-1 in 1890 to over 400 ha-1 by 1970 (Adams 1975). On another site
in central Oregon where trees were absent prior to 1880, J. occidentalis increased to 1018 ha-1 by
1980 (Eddleman 1987). Recent expansion is similar to increases in other Juniperus species
throughout western United States (Ellis and Schuster 1968, Tausch et al. 1981, West 1984,
Tausch and West 1988).
The objectives of our study were to (1) describe the chronology of J. occidentalis expansion
during the past several centuries in southeastern Oregon, (2) determine the effect of plant canopy
and interspace on J. occidentalis seedling establishment and growth rates, and (3) determine the
age when J. occidentalis reaches maximum reproductive potential.
Return to Contents
Study Area
The study area is located on Steens Mountain in southeastern Oregon, approximately 80 km south
of Burns. This isolated volcanic fault-block, which lies in the extreme northwest Basin and Range
Province (Fenneman 1931), is about 80 km long and oriented in a northeast direction (Baldwin
1981). The elevation of Steens Mountain ranges from 1268 to 2949 m, with a steep east-facing
escarpment and a gentle west-facing slope. Climate is cool and semiarid, characteristic of the
northern Great Basin. Annual precipitation at the lower elevations averages 220-280 mm,
increasing to greater than or equal to 700 mm at higher elevations (NOAA 1993). Most moisture is received as snow in
November, December, and January and as rain in March through June.
J. occidentalis woodlands on Steens Mountain form a discontinuous belt between 1450 and 2100
m in elevation. Severe winter conditions probably restrict J. occidentalis from expanding into
higher elevations (Billings 1954, Mehringer 1987). Limited distribution below 1500 m is possibly
due to a combination of late spring frosts (Billings 1954) and limiting moisture. Tree canopy
cover varies from open to 30% cover, except on mesic P. tremuloides sites where J. occidentalis
cover approaches 100%. However, based on age structure and canopy leader growth, tree
canopies are still actively expanding on the majority of sites measured. Early observations on
Steens Mountain indicate the landscape contained only scattered stands of J. occidentalis
(Griffiths 1902). Since 1900 the abundance of J. occidentalis pollen in the Steens Mountain area
has increased fivefold (Mehringer and Wigand 1990).
Plant communities characteristic of J. occidentalis woodlands are Artemisia tridentata ssp.
vaseyana/Festuca idahoensis (Idaho fescue), Artemisia arbuscula/F. idahoensis, and P.
tremuloides . P. tremuloides communities on Steens Mountain range in elevation from 1760 to
2400 m. At lower elevations, in the J. occidentalis woodland belt, P. tremuloides stands form
long, narrow communities along north aspects, which capture windblown snow and runoff.
Return to Contents
Plot Layout
Plot locations were selected in an attempt to reflect sagebrush-grassland communities in different
stages of J. occidentalis invasion on the west slope of Steens Mountain. Old stands on the rocky
outcrops, which make up only a small percentage of present-day woodlands, were not measured.
Sites selected support, or have the potential to support, sagebrush-grassland communities.
Currently these sites are occupied by varying numbers and sizes of J. occidentalis dominance,
creating a woodland structure of dispersed, intermediate, and closed tree stands (Table 1).
| Closed | Abundant adult trees generally greater than 5m tall and usually several trees greater than 130 yr of age, with little understory, particularly on south slopes |
| Intermediate | Abundant J. occidentalis of all age classes, with a more open tree canopy and an understory beginning to decline; trees greater than 130 yr of age are rare |
| Dispersed | Abundant young trees less than 2m tall, a few adult trees but old trees absent, and a well-developed understory. |
Twenty-two 0.4-ha plots were located within the J. occidentalis belt of Steens Mountain; they ranged from 1500 to 2000 m in elevation and were distributed 32 km along the mountain range. Plots were situated along an elevation gradient representing communities from the lower- to upper-elevation J. occidentalis woodland belt. Dominant understory vegetation in the dispersed and intermediate plots was A. tridentata spp. vaseyana and Festuca idahoensis (13 stands), A. arbuscula and F. idahoensis (4 stands), and a mosaic of A. arbuscula and A. tridentata ssp. vaseyana (2 stands). Understory vegetation in the closed stands (n = 3) comprised a few remnant deep-rooted perennial grasses, skeletons of dead A. tridentata ssp. vaseyana, and 70% bare ground (EOARC data file).
An additional six 0.1-ha plots were established in six separate P. tremuloides stands. Three stands
were in advanced stages of J. occidentalis invasion with few to no adult P. treinuloides trees and
dead P. tremuloides trunks on the ground. The remaining three stands were characterized by a
dominant P. tremuloides overstory and an understory of young J. occidentalis. Elevation for the
J. occidentalis-P. tremuloides plots ranged from 1930 to 2000 m, all with a similar northeast
aspect.
Return to Contents
Measurements
Prior to sampling, string was stretched along the contour of each 0.4-ha plot at 1-m intervals to
keep track of measured trees. J. occidentalis density (trees ha-1) was recorded for trees <0.5 m
tall, defined as adult, across the entire plot. Tree height, minimal and maximal crown diameters,
and basal area just above the trunk swell at the stem base near the litter layer were recorded. Tree
height was measured with a tape for trees less than or equal to 2 m and a clinometer for trees >2 m tall. Tree
canopy cover was estimated by adding crown area measurements of all trees for each plot.
Similar measurements were recorded on juvenile trees (defined as trees less than or equal to 0.5 m tall), but only
those on the lower left quarter (0.1 ha) of each 0.4 ha plot. Current-year J. occidentalis seedlings
(any plant with cotyledons still attached) were not recorded. Establishment location of each
juvenile tree was recorded: beneath the canopy of J. occidentalis, Artemisia, other shrubs,
tussock grass, or in the interspace. Less than 1% of juveniles were located beneath other shrubs
or grasses; therefore, only J. occidentalis, Artemisia, and interspace are reported.
J. occidentalis is considered submonoecious (Vasek 1966). Male and female reproductive status
was determined by estimating abundance of cones and berries for each tree. Abundance was
ranked in four classes: (0) absent, (1) scarce, (2) obvious but not abundant, and (3) abundant.
In each plot a 10-tree subsample was randomly selected for aging in each of four height classes:
(1) <0.5 m, (2) 0.5-1.8m, (3) 1.8-3 m, and (4) >3 m. In several of the dispersed plots, sample size
for trees >3 m was smaller than 10, due to a lack of trees. We also sampled all old trees on plots
when they occurred (n = 0-5 ha-1), old trees were easily identified by their growth form,
containing rounded tops and heavy limbs, and lacking strong terminal leader growth (Burkhardt
and Tisdale 1969). A cross section was removed approximately 30 cm above ground level from
each tree >0.5 m tall and at ground level for trees less than or equal to 0.5 m, and then brought back to the lab for
aging. Two radii from each cross section were polished, stained, and counted. Age was
estimated by averaging both radii and adding 10 yr to correct for the 30-cm base. Mean
differences between radii were 4% for trees >50 yr and 1% for trees less than or equal to 50 yr of age. Adams
(1975) reported that growth-ring characteristics of J. occidentalis are useful in
dendrochronological studies. The presence of false and missing rings was similar to that for Pinus
ponderosa. Over 1200 trees were aged and approximately 14,000 counted and measured. In the
six P. tremuloides stands, density of both J. occidentalis and P. tremuloides and age and height
for J. occidentalis were measured across the entire 0.1-ha plot.
Evidence indicated minimal J. occidentalis mortality has occurred on Steens Mountain during the
past 120 years. We observed very few dead or dying trees for all age classes (excluding
seedlings), except where individual J. occidentalis trees had been cut or burned. Mortality of
Juniperus species rapidly declines following the seedling stage (Van Pelt et al. 1990). Juniperus
has few pests that prove fatal to the tree (Lanner 1984). We avoided recently cut or burned
stands, which constituted a small percentage of J. occidentalis- occupied stands. Where remains
of dead trees were observed, we noted they persisted for a long period of time. By recutting
several stumps adjacent to one of our plots and aging and matching ring widths with adjacent live
trees, we determined these trees were harvested around 1920. Others have also observed the
persistence of Juniperus stumps (Young and Budy 1979).
Return to Contents
Statistical Analysis
Height growth data for adult trees were analyzed using a randomized complete block design in
PROC GLM of SAS (SAS 1986). Means were separated using Duncan's Multiple Range Test at
p less than or equal to 0.05 level. A split-plot design was used in the analysis of juvenile height growth. Main plots
were sites and subplots were location of establishment (interspace, Artemisia, J. occidentalis). A
Duncan's Multiple Range Test was used to separate the means.
Return to Contents
Results
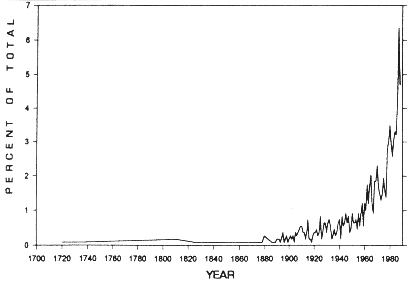
Figure 1. Years of establishment for Juniperus occidentalis trees on Steens Mountain, Oregon (n=1200).
Little change in J. occidentalis density appeared to occur between the early 1700s and the 1880s
(Fig. 1). We encountered old trees (standing trees >130 years old, large stumps, and burned-out
trunks) on several A. arbuscula flats and A. tridentata ssp. vaseyana communities. However,
data indicated presettlement tree densities in these Artemisia communities were less than or equal to 5 trees ha-1,
suggesting very open J. occidentalis stands. The first evidence of an increase in tree densities
occurred in the 1880s, with relatively steady establishment ensuing into the 1950s, similar to that
observed by Tausch and West (1988). In the 1960s J. occidentalis establishment began occurring
at a geometric rate.
| Density (# ha-1) | Density (# ha-1) | Establishment site % (for juveniles) | Establishment site % (for juveniles) | Establishment site % (for juveniles) | ||||
|---|---|---|---|---|---|---|---|---|
| # sites | Canopy cover % | Basal area (m2ha-1) | Adults > 0.5 m ht | Juveniles 0.5m ht | J. occidentalis | Artemisia | Interspace | |
| A. tridentata ssp. vaseyana | ||||||||
| closed | 6 | 22 (18-28) | 5.2 (3.1-9.8) | 296 (217-496) | 580 (118-1226) | 86a1 | 9b | 5b |
| intermediate | 8 | 6 (5-10) | 1.8 (0.5-4.7) | 95 (50-165) | 815 (335-1423) | 29b | 58a | 13c |
| dispersed | 2 | 2 (1-3) | 0.4 (0.2-0.6) | 52 (31-70) | 188 (96-280) | 3b | 50a | 47a |
| A. arbuscula | ||||||||
| closed | 3 | 15 (12-20) | 3.5 (1.8-5.4) | 158 (74-247) | 99 (20-198) | 27b | 67a | 6c |
| intermediate | 3 | 6 (4.5-6.7) | 1.8 (0.9-3.2) | 104 (77-153) | 375 (167-790) | 11b | 61a | 28b |
Closed J. occidentalis stands, which once supported A. tridentata ssp. vaseyana, were
characterized by an abundance of adult trees (greater than or equal to 3 m tall), a tree canopy cover of 18-28% (Table
2), and the presence of a few old trees (130+ yr; 2 to 5 ha-1). J. occidentalis densities began
increasing in these stands between 1878 and 1890. In the intermediate J. occidentalis stands, trees
> 130 yr were rare. Tree canopy cover ranged from about 8 to 16%, and densities of adult trees
varied from 35 to 100 ha-1. Trees <3 m in height, particularly juveniles, were abundant. J.
occidentalis expansion in these sagebrush- grassland communities began between 1890 and 1910.
In the dispersed stands few trees were >60 yrs old, and we aged no trees > 100 yr. Tree canopy
cover was usually <5% in the dispersed stands and densities of large adult trees <35 ha-1. Invasion
of J. occidentalis into these sage brush-grassland communities began after 1930.
| P. tremuloides | P. tremuloides | J. occidentalis | J. occidentalis | |
|---|---|---|---|---|
| Stage of succession | Adult | Juvenile | Adult | Juvenile |
| Late (n=3) | 17 (0-50) | 1316 (0-3952) | 1392 (929-2203) | 9462 (4327-18791) |
| Intermediate (n=3) | 1060 (476-1670) | 6553 (5266-9480) | 1090 (632-1739) | 2816 (622-5968) |
Greatest densities of J. occidentalis trees measured on Steens Mountain occurred in P.
tremuloides sites (Table 3). In the late stages of J. occidentalis succession on these sites, tree
canopy cover approached 100%. Live P. tremuloides occurred only on one of the three sites, and
almost all trees were <0.5 m tall. In the remaining two stands only the remnants of large P.
tremuloides trunks decaying in the understory were present. J. occidentalis invasion in these P.
tremuloides sites began between 1910 and 1920. No J. occidentalis trees >80 yr were
encountered. In stands with a P. tremuloides overstory, P. tremuloides density of small shoots
was greater than that of J. occidentalis. However, P. tremuloides size classes between 0.5 m and
large adults were absent, indicating a lack of P. tremuloides stand rejuvenation. On these sites J.
occidentalis invasion began between 1930 and 1940.
Height growth for young J. occidentalis trees (<20 yr) across all sites averaged 2.9 cm yr-1.
Based on growth rates and height of trees between 10 and 20 yr of age (n = 200) across all
Artemisia sites, 90% of trees 15 yr old were <1 m tall (64% were <0.5 m tall). Surprisingly,
height growth rates of juvenile trees did not significantly differ between A. arbuscula and A.
tridentata ssp. vaseyana communities. However, location of establishment within communities
significantly influenced growth rates of young J. occidentalis trees (Table 4). Trees establishing
beneath an Artemisia canopy grew faster than young trees growing in the interspace.
| Establishment Site | cm yr-1 |
|---|---|
| Artemisia | 3.3 A |
| J. occidentalis | 2.7 AB |
| Interspace | 2.4 B |
Shrub and tree canopies also significantly influenced location of J. occidentalis seedling
establishment in Artemisia communities. The largest proportion of juvenile trees was usually
located beneath canopies of A. tridentata ssp. vaseyana or A. arbuscula and J. occidentalis
(Table 2). Less than 20% of juveniles across all 22 Artemisia sites established in the interspace.
On Steens Mountain, for trees >0.5 in tall, 32% expressed predominantly only male or only female characteristics, 38% both male and female, and 30% contained neither fruits nor
cones. J. occidentalis trees producing abundant crops of cones or berries were either male or
female dominant. No trees were measured which contained an abundant crop of both berries and
cones. Sixty-five percent of J. occidentalis trees with an abundant crop of berries contained no
male cones. The remaining 35% contained only a scarce number of cones. The majority of trees
producing abundant crops of male cones contained only scarce numbers of berries. Approximately
75% of trees producing heavy crops of berries or cones were >50 yr old. Trees <20 yr old
expressing reproductive effort were rare and produced only a few cones or berries. The ratio of
trees producing large crops of cones versus berries (cones:berries) increased from 1.7 in the
scattered J. occidentalis stands to 3.8 in the closed stands.
Return to Contents
Discussion
Low densities and limited distribution of J. occidentalis trees > 130 yr and limited numbers of
dead trees or old stumps suggest J. occidentalis has greatly expanded on Steens Mountain during
the past 100 yr. Distribution of old trees was generally limited to rocky ridges and A. arbuscula
communities. Old trees were found only occasionally growing in deeper, well-drained soils such
as A. tridentata ssp. vaseyana grassland communities and were absent in P. tremuloides
communities. In northeastern California, Barbour and Major (1977) found a similar distribution
of old and young J. occidentalis trees. A. tridentata ssp. vaseyana and A. arbuscula
communities, which contained a low density of J. occidentalis trees prior to settlement, were the
earliest sites to initiate an increase in J. occidentalis. Dates of initial establishment of closed and
intermediate stands were similar to periods of early stand development reported by Young and
Evans (1981) in northeastern California and Eddleman (1987) in central Oregon.
Expansion of J. occidentalis coincides with Euro-American settlement in this portion of the Great
Basin. Although no direct cause-and effect relationship can be drawn, we hypothesize that
climate, altered fire frequencies, and grazing in the late 1800s were primary factors initiating the
recent expansion of J. occidentalis. Following the end of the Little Ice Age in the mid 1800s
(Bryson 1989), winters became more mild and precipitation increased above the present long-term
average in the northern half of the Great Basin between 1850 and 1916 (Antevs 1948, Graumlich
1985). Mild, wet winters and cool, wet springs promote vigorous growth in J. occidentalis
(Earle and Fritts 1986, Fritts and Xiangdig 1986).
Presettlement fire-return intervals in A. tridentata ssp. vaseyana communities have been reported
to vary from 15 to 25 yr (Houston 1973, Burkhardt and Tisdale 1976, Martin and Johnson 1979).
Burkhardt and Tisdale (1976) concluded that fire-frequency intervals of 30-40 yr would be
adequate to keep J. occidentalis from invading a sagebrush-grassland community. Following
settlement, frequency of fire in sagebrush grasslands has greatly declined. The reduction of fine
fuels by high densities of domestic livestock greatly reduced the potential for fire in the
Intermountain Shrub Region (Burkhardt and Tisdale 1976, West 1988). Griffiths' (1902)
observations of the overgrazed landscape on Steens Mountain support this hypothesis. Fires set
by Native Americans also declined in the 19th century due to large reductions in their populations
caused by European diseases (Thompson 1916, Cressman 1981) and relocation to reservations in
the 1870s.
The invasion of conifers into P. tremuloides communities is a common occurrence throughout the
western U.S. However, conifers reported to typically invade P. tremuloides stands are species
adapted to more mesic sites such as Pinus contorta (lodgepole pine), P. ponderosa, Pseudotsuga
menziesii (Douglas-fir), Abies concolor (white fir), Abies lasiocarpa (subalpine fir), Picea
engelmannii (Engelmann spruce), and Picea pungens (blue spruce) (Bartos 1973, Mueggler
1985). Invasion of the more drought-tolerant J. occidentalis into P. tremuloides stands is not
well documented.
P. tremuloides is frequently considered a fire-induced species, replaced by less fire tolerant
conifers (Baker 1925, Daubenmire 1943, Mueggler 1976). Prior to settlement, lightning and
human-set fires probably helped maintain many P. tremuloides communities. However, the
occurrence of fire in P. tremuloides stands in the Rocky Mountains has been greatly reduced since
the late 1800s (Jones and DeByle 1985). Mueggler (1985) suggested the combination of fire
suppression and heavy grazing in P. tremuloides communities may favor the establishment of
conifers.
An increase in Artemisia cover may also enhance the invasion of J. occidentalis. As a sagebrush-grassland community shifts towards a greater dominance of shrubs, the number of safe sites for J.
occidentalis seedling establishment increases. Others have also reported the majority of J.
occidentalis seedlings established beneath Artemisia canopies (Burkhardt and Tisdale 1976,
Eddleman 1987). In west Texas, J. pinchotii frequently establishes beneath mesquite plants
(McPherson et al. 1988). Shading by nurse plants may benefit J. occidentalis seedlings (Johnsen
1962) by reducing summer surface temperatures by 45-57% of bare ground surface temperatures
(Burkhardt and Tisdale 1976). Enhanced growth rates of young trees growing beneath A.
tridentata ssp. vaseyana suggest microclimates beneath shrub canopies are more beneficial than
conditions in the interspace. Burkhardt and Tisdale (1976) reported J. occidentalis seedling
growth rates were correlated positively with Artemisia and correlated negatively with bare
ground.
J. occidentalis approached full reproductive potential near 50 yr. As J. occidentalis densities
increased, the proportion of trees became predominantly male across sites. Highly fecund female
trees appeared to be most important in open stands where J. occidentalis was actively expanding.
In central Oregon, Eddleman (1984) observed that trees in the interior woodlands were strongly
dominated by male cone production while trees growing in the open produced more female cones.
He also reported trees did not produce significant quantities of fruit until 50-70 yr of age.
Return to Contents
Conclusion
Optimal climatic conditions around the turn of the century, reduced fire return intervals, and the
indirect effect of livestock through the reduction of fine fuels and an increase in Artemisia cover
are probably primary factors that have contributed to the rapid expansion of J. occidentalis in
southeast Oregon during the late 1800s and early 1900s. The accelerated increase in J.
occidentalis density and invasion during the last 30 years into new communities is probably
largely due to the continued absence of fire, abundant woody plant cover, and the large increase in
J. occidentalis seed rain.
Return to Contents
Acknowledgements
This is Technical Report 10,494 of the Eastern Oregon Agricultural Research Center, Oregon State University.
Literature Cited
Adams, A.W. 1975. A brief history of juniper and shrub populations in southern Oregon. Wildlife
Research Report 6, Oregon State Wildlife Commission, Corvallis.
Antevs, E. 1948. Climatic changes and pre-white man. Pages 168-191 in The Great Basin, with
emphasis on glacial and postglacial times. University of Utah Biological Service Bulletin 10(7).
Baker, F.S. 1925. Aspen in the central Rocky Mountain region. USDA Bulletin 1291.
Baldwin, E.M. 1981. Geology of Oregon. Kendal/Hunt Publishing Co., Dubuque, IA.
Barbour, M.C. and J. Major. 1977. Terrestrial vegetation of California. Wiley Interscience, New York, NY.
Barney, M.A., and N.C. Frishknecht. 1974. Vegetation changes following fire in the pinyon-juniper type of west-central Utah. Journal of Range Management 74:91-96.
Bartos, D.L. 1973. A dynamic model of aspen succession. Pages 13-25 in Proceedings, IUFRO
Biomass Studies. IUFRO.
Billings, W.D. 1954. Temperature inversions in the pinyon-juniper zone of a Nevada mountain
range. Butler University Botanical Studies 12.
Blackburn, W.H. and P.T. Tueller. 1970. Pinyon and juniper invasion in black sagebrush
communities in east-central Nevada. Ecology 51: 841-848.
Bryson, R.A. 1989. Late Quaternary volcanic modulation of milankovith climate forming.
Theoretical Applied Climatology 39: 115-125.
Burkhardt, J.W, and E.W Tisdale. 1969. Nature and successional status of western juniper
vegetation in Idaho. Journal of Range Management 22: 264-270.
_____. 1976. Causes of juniper invasion in southwestern Idaho. Ecology 76: 472-484.
Caraher, D.L. 1977. The spread of western juniper in central Oregon. Pages 3-8 in R.E. Martin, J.
E. Dealy. and D. L. Caraher, eds., Proceedings, Western juniper Ecology and Management
Workshop. USDA Forest Service, General Technical Report PNW-4.
Cottam, W.P., and G. Stewart. 1940. Plant succession as a result of grazing and of meadow
desiccation by erosion since settlement in 1892. Journal of Forestry 38:613-626.
Cressman, L.S. 1981. The sandal and the cave. Oregon State University Press, Corvallis.
Cronquist, A., A.H. Holmgren, N.H. Holmgren, and J.L. Reveal. 1972. Intermountain flora:
vascular plants of the Intermountain West, U.S.A. Volume 1. Hafner Publishing Company, New
York, NY.
Daubenmire, R.F. 1943. Vegetational zonation in the Rocky Mountains. Botanical Review 9:
325-393.
Dealy, J. E., J.M. Geist, and R.S. Driscoll. 1978. Western juniper communities on rangeland of
the Pacific Northwest. Pages 201-204 in D.E. Hyder, ed., Proceedings, First International
Rangeland Congress, Denver, CO.
Earle, C.J., and H.C. Fritts. 1986. Reconstructing river flow in the Sacramento Basin since 1560.
Report, California Department of Resources, Agreement DWR B-55395. Laboratory of Tree-ring
Research, University of Arizona, Tucson.
Eddleman, L.E. 1984. Ecological studies on western juniper in central Oregon. Pages 27-35 in
T.E. Bedell, ed., Proceedings, Western juniper Management Short Course. Oregon State
University and Extension Service, Corvallis.
____. 1987. Establishment and stand development of western juniper in central Oregon. Pages
255-259 in R.L. Everett, ed., Proceedings, Pinyon-juniper Conference. USDA Forest Service,
General Technical Report INT-215.
Ellis, D., and J.C. Schuster. 1968. juniper age and distribution on an isolated butte in Garza
County, Texas. Southwestern Naturalist 13: 343-348.
Fenneman, N.M. 1931. Physiography of the western United States. McGraw-Hill, New York,
NY.
Franklin, J.F, and C.T. Dyrness. 1973. Natural vegetation of Oregon and Washington. USDA
Forest Service, General Technical Report PNW-8. Portland, OR.
Fritts, H.C., and W. Xiangdig. 1986. A comparison between response-function analysis and other
regression techniques. Tree-ring Bulletin 46: 31-46.
Graumlich, L. 1985. Long-term records of temperature and precipitation in the Pacific Northwest
derived from tree rings. Unpublished doctoral dissertation, University of Washington, Seattle.
Griffiths, D. 1902. Forage conditions on the northern border of the Great Basin. Bureau of Plant
Industry, USDA, Bulletin 15.
Hopkins, W.E. 1979. Plant associations of the Fremont National Forest. USDA Forest Service,
Pacific Northwest Region, R6-ECOL-79-004.
Houston, D.B. 1973. Wildfires in northern Yellowstone National Park. Ecology 54: 1109-1117.
Johnsen, T.N. 1962. One-seed juniper invasion of northern Arizona grasslands. Ecological
Monographs 32: 187-207.
Johnson, C.G., JR., and S.A. Simon. 1987. Plant associations of the Wallowa-Snake Province,
Wallowa-Whitman National Forest. USDA Forest Service, Pacific Northwest Region Report R6-ECOL-TP-255B-86. Portland, OR.
Jones, J.R. and N.V. DeByle. 1985. Fire. Pages 77-81 in N.V. DeByle and R.P. Winokur, eds.,
Aspen: ecology and management in the western United States. USDA Forest Service General
Technical Report RM-119.
Lanner, R.M. 1984. Trees of the Great Basin: a natural history. University of Nevada Press,
Reno.
Martin, R.E., and A.H. Johnson. 1979. Fire management of Lava Beds National Monument.
Pages 1209-1217 in R.M. Linn, ed., Proceedings, First Conference of Science and Research in the
National Parks. USDI National Park Service, Transactions Proceedings Serial 5.
McPherson, G.R., H.A. Wright, and D.B. Wester. 1988. Patterns of shrub invasion in semiarid
Texas grasslands. American Midland Naturalist 102: 391-397.
Mehringer, P.J., Jr. 1987. Late Holocene environments on the northern periphery of the Great
Basin. Final report, Bureau of Land Management, Portland, OR.
Mehringer, P.J., Jr. and P.E. Wigand. 1990. Comparison of late Holocene environments from
woodrat middens and pollen: Diamond Craters, Oregon. Pages 294-325 in J.L. Betancourt, T.R.
Van Devender, and P.S. Martin, eds., Packrat middens: the last 40,000 years of biotic change.
University of Arizona Press, Tucson.
Mitchell, V.L. 1976. The regionalization of climate in the western United States. Journal of
Applied Meteorology 15: 920-927.
Mueggler, W.F. 1976. Type variability and succession in Rocky Mountain aspen. Pages 16-19 in
Proceedings, Utilization and Marketing Tools for Aspen Management in the Rocky Mountains.
USDA Forest Service, General Technical Report RM-29.
____ 1985. Vegetation associations. Pages 45-55 in N.V. DeByle and R.P. Winokur, eds., Aspen:
ecology and management in the western United States. USDA Forest Service, General Technical
Report RM-119.
Neilson, R.P. 1987. On the interface between current ecological studies and the paleobotany of
pinyon-juniper woodlands. Pages 93-98 in R. Everett, ed., Proceedings, Pinyon-juniper
Conference. USDA Forest Service, General Technical Report INT-215.
Nichol, A.A. 1937. The natural vegetation of Arizona. University ofArizona Technical Bulletin
68.
NOAA. 1993. National Climatic Data Center. Federal Building, Asheville, NC.
Rich, E.E., A.M. Johnson, and B.R. Baker, eds. 1950, Peter Skene Ogden's Snake Country
journals: 1824-25 and 1825-26. The Hudson Bay Society, London.
SAS. 1986. SAS-STAT user's guide (release 603). SAS Institute, Inc., Cary, NC.
Tausch, R.J., N.E. West, and A.A. Nabi. 1981. Tree age and dominance patterns in Great Basin
pinyon-juniper woodlands. Journal of Range Management 34:259-264.
Tausch, R J., and N.E. West. 1988. Differential establishment of pinyon and juniper following fire.
American Midland Naturalist 119:174-184.
Thompson, D. 1916. David Thompson's narrative. J.B. Tyrrel, ed. The Champlain Society,
Toronto, Ontario, Canada.
USDI-BLM. 1990. The juniper resources of eastern Oregon. USDI, Bureau of Land Management
Information Bulletin OR-90-166.
Van Pelt, N., R. Stevens, and N.E. West. 1990. Survival and growth of immature Juniperus
osteosperma and Pinus edulis following woodland chaining in central Utah. Southwestern
Naturalist 35: 322-328.
Vasek, F.C. 1966. The distribution and taxonomy of three western junipers. Brittonia 18: 350-372.
West, N.E. 1984. Successional patterns and productivity of pinyon-juniper ecosystems. Pages
1301-1332 in Developing strategies for range management. Westview Press, Boulder, CO.
____1988. Intermountain deserts, shrub steppes, and woodlands. Pages 209-230 in M.B. Barbour
and W.D. Billings, eds., North American terrestrial vegetation. Cambridge University Press,
Cambridge, MA.
Young, J.A., and J.D. Budy. 1979. Historical use of Nevada's pinyon-juniper woodlands. Journal
of Forest History 23: 113-121.
Young, J.A., and R.A. Evans. 1981. Demography and fire history of a western juniper stand.
Journal of Range Management 34: 501-506.
Return to Contents